
Общие свойства молекул вторичных посредников
Внутриклеточные вторичные посредники имеют некоторые общие для них всех свойства:
Вторичные (внутриклеточные) мессенджеры – малые молек или ионы, внутриклет концентрац кот повышает ответ на соединение рецептора с сигнальной молекулой. Роль вторичных мессенджеров выполняют молекулы и ионы: • ц-АМФ (циклический аденозин-3′,5′-монофосфат); • ИФ3 (инозитол-1,4,5-трифосфат); •ДАГ •ц-ГМФ • Са2+.
Аденилатциклазная система включает: — интегральные белки цитоплазматической мембраны: • Rs — рецептор первичного мессенджера – активатора, рецептор первичного мессенджера — ингибитора АЦС; • фермент аденилатциклазу (АЦ). — «заякоренные» белки: • Gs — ГТФ-связывающие белки • цитозольный фермент протеинкиназу А (ПКА).
Рис. 4.11. Функционирование аденилатциклазной системы
Когда соответствующий лиганд (например, адреналин) взаимодействует с рецептором, происходяi конформационные изменения рецепторного белка. Структурно перестроенный G-белок активизирует аденилатциклазу — фермент, располагающийся в мембране. Активизированная аденилатциклаза катализирует превращение АТФ в ц-АМФ. В системе ц-АМФ, внеклеточный сигнал опосредован через белок G, который активирует фермент фосфодиэстеразу. При участии этого фермента из фосфолипида внутреннего слоя мембраны образуется инозитолтрифосфат, который диффундирует в цитоплазму. Здесь он вызывает выброс Са2+ из эндоплазматической сети. Повышение концентрации Са2+ в цитоплазме активирует протеинкиназу, которая фосфорилирует и, следовательно, активирует другие ферменты, участвующ во многих метаболических процессах в клетке. Участие аденилатциклазной системы в регуляции экспрессии генов. Многие белковые гормоны: глюкагон, вазопрессин, паратгормон и др. могут не только изменять скорости реакций путем фосфорилирования ферментов, но и увеличивать или уменьшать их количество, регулируя экспрессию генов.
Вторичные посредники, или «вторичные мессенджеры» — это внутриклеточные сигнальные молекулы, высвобождаемые в ответ на стимуляцию рецепторов и вызывающие активацию первичных эффекторных белков. Они же, в свою очередь, приводят к активации вторичных эффекторных белков. Это запускает каскад физиологических изменений, которые могут быть важны для обеспечения таких важных физиологических процессов, как рост, развитие и дифференцировка клеток, активация деления клетки, транскрипция или, наоборот, угнетение транскрипции генов, биосинтез белков, секреция гормонов, нейромедиаторов или цитокинов соответственно типу клетки, изменение биоэлектрической активности клетки, миграция клеток, обеспечение их выживаемости или, наоборот, индукция апоптоза. Вторичные посредники являются инициирующими элементами во множестве внутриклеточных сигнальных каскадов. Вследствие всего этого вторичные посредники играют очень важную роль в жизни клетки, а грубое нарушение работы любой из систем вторичных посредников оказывает неблагоприятное воздействие на клетку (например, может привести к её опухолевой трансформации или, наоборот, к апоптозу).
Вторичные посредники могут быть синтезированы и активированы при помощи различных ферментов, например, циклаз (аденилатциклаза, гуанилатциклаза), которые синтезируют циклические нуклеотиды цАМФ и цГМФ, или синтазы оксида азота, которая синтезирует оксид азота (II), или фосфолипазы C, которая синтезирует диацилглицерин и инозитолтрифосфат, или же при помощи открытия или закрытия ионных каналов клетки, которые позволяют вход в клетку или выход из клетки тех ионов, как это происходит при передаче сигнала при помощи ионов кальция. Эти малые молекулы вторичных посредников связываются со вторичными эффекторными белками, в частности, активируют протеинкиназы, открывают и закрывают ионные каналы и т. д., что в свою очередь запускает дальнейший внутриклеточный сигнальный каскад вплоть до эффекторов N-го порядка.
Текущая версия страницы пока не проверялась опытными участниками и может значительно отличаться от версии, проверенной 15 октября 2018 года; проверки требуют 8 правок.
У этого термина существуют и другие значения, см. Трансдукция.
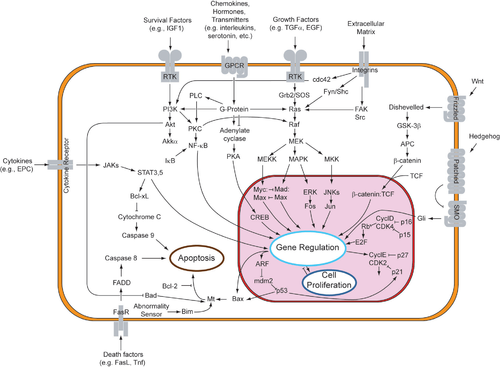
Наиболее хорошо изученные сигнальные пути: PI3K-Akt, JAK-STAT, NF-kB, гетеротримерных G-белков (в том числе аденилатциклазный), Ras-MAPK/ERK, Wnt, Hedgehog, Fas.
Передача сигнала (сигнальная трансдукция, трансдукция, сигналинг, сигнализация, англ. ) — в молекулярной биологии термин «Передача сигнала» относится к любому процессу, при помощи которого клетка превращает один тип сигнала или стимула в другой.
Существование сложных многоклеточных организмов возможно благодаря координации биохимических процессов, протекающих в их клетках. Основой такой координации служат межклеточная коммуникация и передача сигнала внутри отдельных клеток. Вместе это даёт возможность одной клетке контролировать поведение остальных.
В большинстве случаев передача сигнала внутри клетки представляет собой цепь последовательных биохимических реакций, осуществляемых ферментами, часть из которых активируется вторичными посредниками. Такие процессы обычно являются быстрыми: их продолжительность — порядка миллисекунд в случае ионных каналов и минут — в случае активации протеинкиназ и липид-опосредованных киназ. Однако в некоторых случаях от получения клеткой сигнала до ответа на него могут проходить часы и даже сутки (в случае экспрессии генов). Пути передачи сигнала, или сигнальные пути, часто бывают организованы как сигнальные каскады (англ. ): количество молекул белков и других веществ, принимающих участие в передаче сигнала, возрастает на каждом последующем этапе по мере удаления от первоначального стимула. Таким образом, даже относительно слабый стимул может вызывать значительный ответ. Это явление называется амплификацией сигнала. Оригинальный термин англ. впервые появился в реферируемых журналах в 1974 году, а в названии статьи фигурировал в 1979 году.
1.взаимодействие внешнего агента (стимула) с клеточным рецептором,
2.активация эффекторной молекулы, находящейся в мембране и отвечающей за генерацию вторичныхмессенджеров,
3. образование вторичныхмессенджеров,
4.активация мессенджерами белков-мишеней, вызывающих генерацию следующих мессенджеров,
Вторичные мессенджеры — это внутриклеточные вещества, концентрация которых строго контролируется гормонами-нейромедиаторами и другими внеклеточными сигналами — первичными мессенджерами (firstmessengers).
В качестве агониста рецептора клетка может использовать специально синтезированные соединения пептидной природы или использовать свои внутриклеточные метаболиты, которые отсутствуют в экстраклеточной среде. Кофермент АТР и глутамат, действующие экстраклеточно, являются мощными нейротрансмиттерами. Природные экстраклеточные лиганды, которые взаимодействуют с рецепторами и активируют их, называют первичными мессенджерами. Они могут быть подразделены на гормоны, нейротрансмиттеры, цитокины, лимфокины, факторы роста, липофильные молекулы имеющие рецепторы, хемоаттрактанты и.т.д. Каждый из этих терминов представляет класс агентов, действующих достаточно специфично. Тем не менее, существуют примеры многофункциональности первичных мессенджеров: АТР и глутамат являются нейротрансмиттерами, когда они секретируются в синапсах. Гормоны пищеварительного тракта, такие как гастрин, холецитокинин и секретин в центральной нервной системе осуществляют многообразные функции нейромодуляторов, влияя на высвобождение других нейротрансмиттеров. Соматостатин, идентифицированный первоначально как агент гипоталамуса, подавляющий секрецию гормона роста, также функционирует в центральной нервной системе как нейротрансмиттер и нейромодулятор. Более того, он является паракринным агентом для клеток поджелудочной железы и гормоном для печени. Фактор роста тромбоцитов TGFb действует также как хемоатрактант и как ингибитор роста. Тромбин является фактором роста, но также вовлекается в свертывание крови как активатор функции тромбоцитов.
Механизмы, преобразующие внешние сигналы во внутриклеточные, находятся в плазматической мембране клетки. Восприятие клетками внешних сигналов в основном происходит благодаря взаимодействию внешних факторов с клеточными рецепторами, расположенными на наружной мембране клеток. Рецепторы распознают и связывают внешний сигнал (сигнальный агент — лиганд) и приводят в дейcтвие внутриклеточные пути передачи информации, ведущие к запуску и регуляции различных внутриклеточных процессов.
Внешний сигнальный агент, называемый первичным посредником, как правило, не проникает внутрь клетки, а специфически взаимодействует с рецепторами наружной клеточной мембраны. В качестве первичных посредников выступают различные химические соединения (гормоны, нейромедиаторы, газотрансмиттеры и др.) или физические факторы (квант света). Однако существуют гидрофобные гормоны (стероидные и тиреоидные), которые способны проникать внутрь клетки, преодолевая при этом липидный бислой, и взаимодействовать там с растворимыми рецепторными белками. Таким же механизмом действия обладают малые молекулы — NO и CO.
Если внешняя сигнальная молекула воздействует на рецепторы клеточной мембраны и активирует их, то последние передают полученную информацию на систему белковых компонентов мембраны, называемую каскадом передачи сигнала. Мембранные белки каскада передачи сигнала подразделяют на:
белки-преобразователи, связанные с рецепторами
ферменты-усилители, связанные с белками-преобразователями (активируют вторичные внутриклеточные посредники, переносящие информацию внутрь клетки).
Так действуют рецепторы, сопряженные с G-белками. Другие рецепторы (ионные каналы, рецепторы с протеинкиназной активностью) сами служат умножителями.
Клеточный цикл его периоды
Клеточный цикл — это период существования клетки от момента её образования путем деления материнской клетки до собственного деления.
Таким образом клеточный цикл составляют два периода:
1) период клеточного роста, называемый » интерфаза «, и
2) период клеточного деления, называемый » фаза М » В свою очередь, в каждом периоде выделяют несколько фаз
Обычно интерфаза занимает не меньше 90% времени всего клеточного цикла. Например, у быстро делящихся клеток высших эукариот последовательные деления происходят один раз в 16-24 часа, и каждая фаза М длится 1-2 часа. Большая часть компонентов клетки синтезируется на протяжении всей интерфазы, это затрудняет выделение в ней отдельных стадий
В интерфазе выделяют фазу G1, фазу S и фазу G2. Период интерфазы, когда происходит репликация ДНК клеточного ядра, был назван » фаза S » Период между фазой М и началом фазы S обозначен как фаза G1, а период между концом фазы S и последующей фазой М — как фаза G2.
Период клеточного деления (фаза М) включает две стадии: митоз (деление клеточного ядра) и цитокинез (деление цитоплазмы). В свою очередь, митоз делится на пять стадий Описание клеточного деления базируется на данных световой микроскопии в сочетании с микрокиносъемкой и на результатах световой и электронной микроскопии фиксированных и окрашенных клеток.
Общие механизмы систем вторичных посредников
Схема работы механизма системы вторичных посредников
В каждой клетке существует несколько систем вторичных посредников, в частности система циклического АМФ, фосфатидилинозитоловая система (диацилглицерин и инозитолтрифосфат), система арахидоновой кислоты, система оксида азота, система кальция, система циклического ГМФ и другие, однако все они очень похожи по общему устройству механизма, несмотря на то, что вовлечённые в реализацию их эффектов эндогенные вещества и общие эффекты их активации могут очень различаться.
В большинстве случаев всё начинается с того, что тот или иной внеклеточный лиганд-агонист связывается со специфическим сайтом (доменом связывания лиганда) на трансмембранном рецепторном белке. Связывание лиганда с этим сайтом рецептора приводит к изменению его пространственной конфигурации. Это изменение пространственной конфигурации рецептора может привести к изменению его активности и к запуску того или иного первичного эффектора. Результатом действия первичного эффектора является синтез и выделение (или, наоборот, угнетение синтеза) тех или иных вторичных посредников, то есть генерация вторичного сигнала (вторичного месседжа, будь то положительный месседж — синтез вторичного посредника, или отрицательный месседж — угнетение синтеза вторичного посредника).
Например, в случае с G-белок-связанными рецепторами, изменение конформации белка-рецептора, происходящее при связывании с ним лиганда, приводит к тому, что специфический внутриклеточный домен рецептора приобретает способность связываться с G-белком. G-белок называется так по ГДФ и ГТФ, которые этот белок способен связывать. G-белок гидрофобен, поэтому он связан с внутренней мембраной клетки, что облегчает связывание G-белка с трансмембранными рецепторами. G-белок является передатчиком и трансформатором сигнала от рецептора внутрь клетки; он состоит из трёх субъединиц: α, β и γ.
Когда G-белок связывается с активированным рецептором, он приобретает способность к обмену ГДФ (гуанозиндифосфата), связанного с его α-субъединицей, на ГТФ (гуанозинтрифосфат). После того, как этот обмен произойдёт, α-субъединица G-белка приобретает способность диссоциироваться от β- и γ-субъединиц. Все три субъединицы при этом остаются мембранно-связанными. После того как α-субъединица диссоциирует от β- и γ-субъединиц и приобретает способность диффундировать вдоль внутреннего слоя клеточной мембраны, α-субъединица, в конце концов, сталкивается в процессе этого движения с другим мембранным белком — так называемым «первичным эффектором», которым в случае G-белка обычно является аденилатциклаза.
В результате этого взаимодействия α-субъединицы G-белка с первичным эффектором (например, с аденилатциклазой), ферментативная активность первичного эффекторного белка изменяется тем или иным образом — в зависимости от типа α-субъединицы (ингибиторной или стимулирующей). В результате первичный эффекторный белок производит вторичный сигнал, которым является, соответственно, уменьшение или увеличение продукции того или иного вещества из субстрата его ферментативной активности. Для аденилатциклазы субстратом является АТФ, а конечным продуктом реакции — циклический АМФ. Для первичного эффектора — фосфолипазы C — субстратом является фосфатидилинозитол, а продуктами реакции — диацилглицерин и инозитолтрифосфат. Для синтазы оксида азота субстратом является L-аргинин, а продуктом реакции — оксид азота (II). Эти продукты реакций и являются вторичными посредниками (вторичными мессенджерами), в то время как лиганд-агонист рецептора называется первичным сигналом или первичным мессенджером. Далее вторичный посредник (или вторичные посредники, если их несколько) могут диффундировать в цитоплазму клетки или в межмембранные пространства (в зависимости от гидрофильности или липофильности), тем самым эффективно передавая многократно усиленный сигнал внутрь клетки. В клетке эти вторичные посредники, в свою очередь, активируют различные так называемые «вторичные эффекторы» (например, в случае цАМФ это прежде всего цАМФ-зависимая протеинкиназа А, в случае инозитолтрифосфата — прежде всего протеинкиназа C; мишеней для воздействия может быть несколько или даже много). А вторичные эффекторы запускают дальнейший каскад нисходящих эффекторных реакций. Конечные эффекты зависят как от типа рецептора и G-белка (стимулирующий или ингибирующий), так и от конкретной системы вторичных посредников и вторичных эффекторов.
В то же время β- и γ-субъединицы G-белка способны активировать или угнетать (в зависимости от типа G-белка) добавочные сигнальные пути, например, GIRK (входящие однонаправленные каналы ионов калия) или фосфолипазу C или синтазу оксида азота. Таким образом, один и тот же рецептор способен активировать сразу несколько систем вторичных посредников и производить «кросс-общение» с сигнальными путями, считающимися более характерными для других типов рецепторов.
Ионы кальция являются одним из важных типов вторичных посредников, которые ответственны за осуществление множества важных физиологических функций, таких, как сокращение мышц, оплодотворение, выделение нейромедиаторов. Ионы кальция в норме связаны со специальными белками и/или хранятся в специальных внутриклеточных компартментах (в частности, в гладком эндоплазматическом ретикулуме), откуда могут быть быстро высвобождены в процессе передачи сигнала. Фермент фосфолипаза C (первичный эффектор) продуцирует вторичные посредники диацилглицерин и инозитолтрифосфат, что приводит к повышению проницаемости каналов ионов кальция. Активный G-белок открывает кальциевые каналы клетки, что приводит к входящему току ионов кальция. Другой вторичный посредник, также являющийся продуктом фосфолипазы C, диацилглицерин, активирует протеинкиназу C, которая способствует активации аденилатциклазы и накоплению цАМФ (ещё одного вторичного посредника).
Жизнь любой клетки,
включая глобальные процессы ее роста,
деления и даже гибели, зависит от внешних
регуляторных сигналов, которые она
воспринимает. Такими сигналами могут
быть физические воздействия (температура,
ионизирующее и другое электромагнитное
излучение) или многочисленные химические
соединения. Хорошо изученными веществами,
которые организм использует для регуляции
жизнедеятельности клеток, являются,
например стероидные гормоны, цитокины
или факторы роста, которые, достигая
клеток-мишеней, вызывают в них специфические
метаболические изменения, связанные в
том числе и с изменением экспрессии
больших групп генов. Не менее сильный
и часто также специфический ответ
вызывают различные физиологически
активные вещества экзогенного
происхождения, например феромоны или
токсины.
Все эти сигналы,
передающиеся через соответствующие
сигнальные молекулы, являются первичными
по отношению к тем каскадам биохимических
реакций, которые запускаются в клетках
в ответ на их воздействие. Первичные
сигналы распознаются клетками благодаря
наличию у них специальных молекул-рецепторов
белковой природы, взаимодействующих с
первичными сигнальными молекулами или
воздействиями физической природы.
Первичный сигнал, как правило, не
действует прямо на те метаболические
процессы в клетке, для регуляции которых
он предназначен. Вместо этого воспринимающий
его рецептор инициирует образование в
клетке промежуточных химических
соединений, запускающих внутриклеточные
процессы, воздействие на которые было
целью первичного внеклеточного сигнала.
Поскольку такие промежуточные соединения
несут в себе информацию о первичном
регуляторном сигнале и являются
вторичными его переносчиками, они
получили название вторичных мессенджеров.
Ими могут быть различные ионы, циклические
нуклеотиды, продукты деградации липидов
и целый ряд других химических соединений
биогенного происхождения.
Использование
эукариотами системы вторичных мессенджеров
переводит их на новый уровень интеграции
всех метаболических и катаболических
процессов, что необходимо для существования
многоклеточных организмов. В частности,
вторичные мессенджеры позволяют
многократно усиливать первичный
регуляторный сигнал от внеклеточных
регуляторных молекул, которые благодаря
этому осуществляют свое действие,
находясь в небольших концентрациях во
внеклеточном пространстве. Кроме того,
многие группы клеток и тканей приобретают
способность к однотипной и одновременной
реакции на первичный регуляторный
сигнал, например на действие гормона
какого-либо органа эндокринной системы.
Это обеспечивает возможность быстрой
адаптации многоклеточного организма
к изменяющимся условиям внутренней и
окружающей среды.
Трансмембранный
перенос первичных сигналов. Для
того чтобы первичный регуляторный
сигнал достиг ядра и оказал свое
воздействие на экспрессию генов-мишеней,
он должен пройти через двухслойную
мембрану именно тех клеток, которым он
предназначен. Как правило, это достигается
благодаря наличию на поверхности клеток
рецепторов белковой природы, специфически
выбирающих из окружающей среды сигналы,
распознать которые они в состоянии
(рис. I.22). В простейшем случае, когда
в качестве низкомолекулярных регуляторов
выступают гидрофобные химические
соединения, растворимые в липидах
мембран (например стероидные гормоны),
для их переноса не используются рецепторы,
и они проникают в клетку путем радиальной
диффузии. Внутри клеток такие соединения
специфически взаимодействуют с белковыми
рецепторами, а образующийся комплекс
переносится в ядро, где оказывает свое
регуляторное воздействие на транскрипцию
соответствующих генов (см. рис. I.22,а).
В
отличие от этого рецепторы мембран,
ориентированные во внеклеточное
пространство, обладают способностью
осуществлять транспорт лиганда-регулятора
внутрь клеток посредством эндоцитоза
(поглощения путем втягивания мембраны)
комплекса лиганд-рецептор в составе
мембранных везикул. Такой механизм
используется, в частности, для переноса
внутрь клеток молекул холестерина,
ассоциированных с рецепторами
липопротеинов низкой плотности (см.
рис. I.22,б).
Другой тип рецепторов, ориентированных
на внеклеточные лиганды, – это
трансмембранные молекулы или группа
молекул. Взаимодействие с лигандом
внешней части таких молекул сопровождается
индукцией ферментативной активности,
ассоциированной с внутриклеточной
частью того же самого полипептида (см.
рис. I.22,в).
Примерами подобных рецепторов, обладающих
активностью тирозиновых протеинкиназ,
являются рецепторы инсулина, эпидермального
фактора роста или фактора роста
тромбоцитов. В синапсах нейронов и
местах контакта нейромышечных тканей
лиганды-нейромедиаторы (например
ацетилхолин или -аминомасляная
кислота) взаимодействуют с трансмембранными
ионными каналами (см. рис. I.22,г).
В ответ на это происходит открытие
ионных каналов, сопровождаемое
перемещением ионов через мембрану и
быстрым изменением трансмембранного
электрического потенциала. Другие
трансмембранные рецепторы осуществляют
связь белков внеклеточного матрикса с
микрофиламентами цитоскелета клеток
и регуляцию формы клеток, зависящую от
внеклеточного матрикса, их подвижности
и роста (см. рис. I.22,д).
Наконец, большая группа внеклеточных
сигналов распознается рецепторами,
ассоциированными на внутренней
поверхности мембраны с GTP-связывающими
белками, которые, в свою очередь, в ответ
на первичный сигнал начинают синтез
вторичных мессенджеров, регулирующих
активность внутриклеточных белков (см.
рис. I.22,е).
Классификация по структурному признаку
рецепторов, осуществляющих перенос
сигнала в клетки через мембраны, приведена
в табл. I.12.
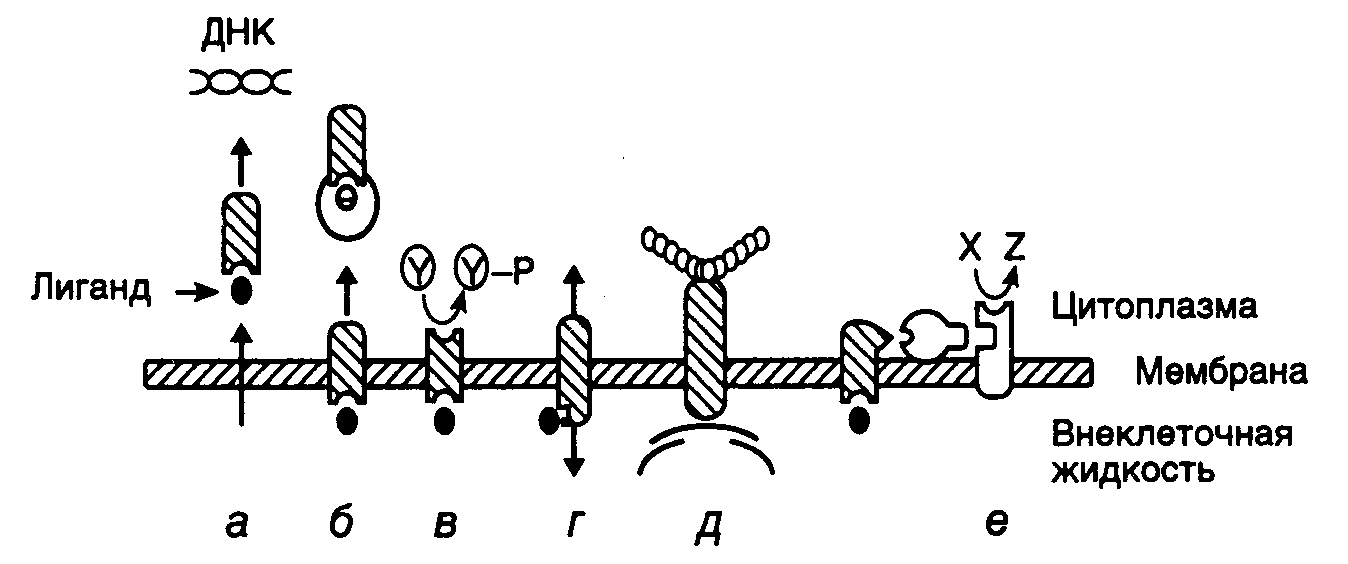
Рис. I.22. Способы передачи
внеклеточных регуляторных сигналов
через мембраны эукариотических клеток
(а–е)
Y и Y–P – нефосфорилированные и
фосфорилированные остатки Tyr в белках
соответственно. Показано также превращение
предшественника X во вторичный мессенджер
Z
Все рецепторы,
участвующие в трансмембранной передаче
сигнала, подразделяют на три класса.
При этом, как правило, учитывается
сходство или различие вторичных структур
субъединиц, а не особенности их
аминокислотных последовательностей.
Рецепторы 1-го класса образуют олигомерные
структуры вокруг пор в мембранах. Перенос
сигнала в этом случае происходит в
результате открытия или (в одном случае)
закрытия ионных каналов. Основная часть
рецепторов 2-го класса погружена в
мембраны, и каждая из субъединиц содержит
последовательности, распознаваемые
G-белками (см. ниже). Для всех субъединиц
этого класса характерно наличие
трансмембранной (ТМ) последовательности,
которая 7 раз пересекает мембрану.
Субъединицы рецепторов 3-го класса
минимально погружены в мембраны, что
обеспечивает подвижность рецепторов
и возможность их интернализации (перехода
в цитоплазму клеток в составе мембранной
везикулы). Большая часть полипептидных
цепей этих субъединиц экспонирована
наружу клеток.
Вторичные
мессенджеры.
Гипотеза о том, что действие гормонов
на метаболизм клеток и экспрессию генов
опосредуется внутриклеточными вторичными
мессенджерами, впервые появилась после
открытия в конце 1950-х годов Е. Сазерлендом
циклического аденозин-3’,5’-монофосфата
(cAMP). К настоящему времени список вторичных
мессенджеров расширился и включает
циклический гуанозин-3’,5’-монофосфат,
фосфоинозитиды, ионы Ca2+
и H+,
метаболиты ретиноевой и арахидоновой
кислот, закись азота (NO), а также некоторые
другие химические соединения биогенного
происхождения. Подробное рассмотрение
особенностей механизма действия каждого
из них выходит за рамки данной монографии,
хотя все они могут оказывать специфическое
влияние на экспрессию генов.
Фармакодина́мика
— раздел фармакологии, изучающий
биохимические эффекты и физиологические
действия лекарств на тело человека, на
микроорганизмы или паразитов, находящихся
внутри тела человека или снаружи. Она
также изучает механизмы действия
лекарств, связь между концентрацией
лекарственных веществ и достигнутым
ими действием.
Многие
лекарства имеют одинаковый механизм
действия
и, следовательно, могут быть объединены
в группы и подгруппы. Количество различных
фармакологических групп (подгрупп)
ограничивается десятками.
Под
действием лекарств в организме не
происходит новых биохимических реакций
или физиологических процессов. Большинство
лекарств только стимулируют, имитируют,
угнетают или полностью блокируют
действие внутренних посредников,
передающих сигналы между различными
органами и системами через биологические
субстраты.
Каждое
звено механизма обратной связи участвует
в регулировании функций клетки и целого
организма, а, следовательно, может
служить “мишенью” – биологическим
субстратом – для лекарственных средств.
Многие
ферменты являются “мишенями” для
лекарств. Лекарства могут угнетать или
– реже – повышать активность этих
ферментов, а также являться для них
“ложными” субстратами. Например,
угнетающими активность (ингибирующими)
ферментов средствами являются
ненаркотические анальгетики и нестероидные
противовоспалительные средства,
некоторые противоопухолевые препараты
(метотрексат), а ложным субстратом –
метилдофа. Ингибиторы ангиотензинпревращающего
фермента (АПФ) (каптоприл и эналаприл)
широко применяются в качестве понижающих
артериальное давление (гипотензивных)
средств. Изменяя активность ферментов,
лекарства изменяют внутриклеточные
процессы и тем самым обеспечивают
лечебный эффект.
В
основе фармакологического действия
лекарств лежит их физико-химическое
или химическое взаимодействие с такими
“мишенями”. Возможность взаимодействия
лекарства с биологическим субстратом
зависит в первую очередь от химического
строения каждого из них. Последовательность
расположения атомов, пространственная
конфигурация молекулы, величина и
расположение зарядов, подвижность
фрагментов молекулы относительно друг
друга влияют на прочность связи и, тем
самым, на силу и продолжительность
фармакологического действия. Молекула
лекарственного вещества в большинстве
случаев имеет очень маленький размер
по сравнению с биологическими субстратами,
поэтому она может соединяться только
с небольшим фрагментом макромолекулы
рецептора. При любой реакции между
лекарством и биологическим субстратом
образуется химическая связь
Итак,
есть сигнальные молекулы (медиаторы,
гормоны, эндогенные биологически
активные вещества), и есть биологические
субстраты, с которыми эти молекулы
взаимодействуют. Лекарства, введенные
в организм, могут воспроизводить или
блокировать эффекты естественных
сигнальных молекул, изменяя тем самым
функции клеток, тканей, органов и систем
органов. Этим определяется фармакологическое
действие лекарств.
Воспроизведение
действия (миметический эффект)
наблюдается в тех случаях, когда молекула
лекарственного вещества и естественная
сигнальная молекула очень похожи: имеют
высокое соответствие физико-химических
свойств и структуры, обеспечивающих
одинаковые внутриклеточные изменения.
Результатом взаимодействия лекарства
с рецептором в этом случае является
активация или торможение определенной
функции клеток в полном соответствии
с действием эндогенной (внутренней)
сигнальной молекулы. Подобным образом
действуют очень многие аналоги гормонов
и медиаторов. Цель создания подобных
лекарств – получение препаратов с более
выраженным, стабильным и длительным по
сравнению с медиатором (адреналин,
ацетилхолин, серотонин и другие)
действием, а также восполнение дефицита
медиатора или гормона и, соответственно,
их функций.
Конкурентное
действие (блокирующий, литический
эффект)
встречается часто и присуще лекарствам,
которые лишь частично похожи на сигнальную
молекулу (например, медиатор). В этом
случае лекарство способно связываться
с одним из участков рецептора, но оно
не вызывает комплекса реакций,
сопутствующих действию естественного
медиатора. Такое лекарство как бы создает
над рецептором защитный экран, препятствуя
его взаимодействию с естественным
медиатором, гормоном и так далее.
Конкурентная борьба за рецептор,
называемая антагонизмом (отсюда и
название лекарств – антагонисты),
позволяет корректировать физиологические
и патологические реакции. Подобным
образом действуют адрено-, холино- и
гистаминолитики.
Следующий
тип взаимодействия лекарства с рецептором
называют неконкурентным,
и в этом случае молекула лекарства
связывается с рецепторной макромолекулой
не в месте ее взаимодействия с медиатором,
а на рядом расположенном участке, то
есть действует опосредованно. При этом
происходит изменение пространственной
структуры рецептора, вызывающее раскрытие
или закрытие его для естественного
медиатора. В этих случаях рецептор для
лекарства и рецептор для медиатора не
совпадают, но находятся в одном рецепторном
комплексе, и лекарство не вступает в
прямое взаимодействие с рецептором.
Ярким примером лекарств, действующих
по этому типу, являются бензодиазепины
– большая группа структурно родственных
соединений, обладающих анксиолитическими,
снотворными и противосудорожными
свойствами. Соединяясь со специфическими
бензодиазепиновыми рецепторами, которые
взаимосвязаны с рецепторами
гамма-аминомасляной кислоты (ГАМК),
лекарственное средство изменяет
пространственную конфигурацию
ГАМК-рецепторов и увеличивает прочность
их связи с субстратом – гамма-аминомасляной
кислотой. В результате усиливается
тормозящее влияние этого медиатора на
центральную нервную систему, чем
обеспечивается лечебный эффект
препаратов.
В
качестве «мишеней» для лекарственных
средств служат рецепторы, ионные каналы,
ферменты, транспортные системы и гены.
Рецепторами называют активные группировки
макромолекул субстратов, с которыми
взаимодействует вещество. Рецепторы,
обеспечивающие проявление действия
веществ, называют специфическими.
Выделяют
следующие 4 типа рецепторов
I.
Рецепторы, осуществляющие прямой
контроль за функцией ионных каналов. К
этому типу рецепторов, непосредственно
сопряженных с ионными каналами, относятся
н-холинорецепторы, ГАМКА-рецепторы,
глутаматные рецепторы.
II.
Рецепторы, сопряженные с эффектором
через систему «G-белки — вторичные
передатчики» или «G-белки-ионные каналы».
Такие рецепторы имеются для многих
гормонов и медиаторов (м-холинорецепторы,
адренорецепторы).
III.
Рецепторы, осуществляющие прямой
контроль функции эффекторного фермента.
Они непосредственно связаны с
тирозинкиназой и регулируют фосфорилирование
белков. По такому принципу устроены
рецепторы инсулина, ряда факторов роста.
IV.
Рецепторы, контролирующие транскрипцию
ДНК. В отличие от мембранных рецепторов
I-III типов, это внутриклеточные рецепторы
(растворимые цитозольные или ядерные
белки). С такими рецепторами взаимодействуют
стероидные и тиреоидные гормоны.
Вторичные
посредники, или «вторичные мессенджеры»
— это внутриклеточные сигнальные
молекулы, высвобождаемые в тех или иных
внутриклеточных сигнальных каскадах
в ответ на стимуляцию тех или иных
рецепторов и вызванную ею активацию
первичных эффекторных белков. Вторичные
посредники, в свою очередь, приводят к
активации вторичных эффекторных белков.
Это, в свою очередь, запускает каскад
тех или иных физиологических изменений,
которые могут быть важны для обеспечения
таких важных физиологических процессов,
как рост, развитие и дифференцировка
клеток, активация деления клетки,
транскрипция или наоборот угнетение
транскрипции тех или иных генов, биосинтез
тех или иных белков, выделение ею
гормонов, нейромедиаторов или цитокинов
соответственно типу клетки, изменение
биоэлектрической активности клетки,
миграция клеток, обеспечение их
выживаемости или, наоборот, индукция
апоптоза. Вторичные посредники являются
инициирующими элементами во множестве
внутриклеточных сигнальных каскадов.
Вследствие всего этого вторичные
посредники играют очень важную роль в
жизни клетки, а грубое нарушение работы
любой из систем вторичных посредников
оказывает неблагоприятное воздействие
на клетку (например, может привести к
её опухолевой трансформации или наоборот
к апоптозу).
Вторичные
посредники, как правило, являются малыми
небелковыми молекулами. Важнейшие
примеры молекул вторичных посредников
(но не ограничивающиеся ими) включают
в себя циклический АМФ, циклический
ГМФ, инозитолтрифосфат, диацилглицерин,
кальций, оксид азота (II).
Соседние файлы в предмете Фармакология
Типы молекул вторичных посредников
Путь передачи внутриклеточного сигнала
Существуют три основных типа молекул вторичных посредников:
Получение клеткой сигнала от первичных посредников обеспечивается особыми белками-рецепторами, для которых первичные посредники являются лигандами. Для обеспечения рецепторной функции молекулы белков должны отвечать ряду требований:
Клеточные рецепторы делятся на следующие классы:
Механизмы активации рецепторов
Ключевые компоненты сигнального пути Ras-MAPK/ERK
Иногда активация рецептора внешним стимулом сразу приводит к ответу клетки. Например, когда нейротрансмиттер ГАМК активирует свой рецептор, входящий в состав ионного канала на поверхности нейрона, канал начинает пропускать ионы хлорида, что приводит к изменению мембранного потенциала всей клетки. В других случаях активация рецептора лишь инициирует цепь событий, передающих регуляторный стимул внутри клетки через более или менее длинную цепь посредников. Такая цепь называется сигнальным путём.
Передача сигнала предполагает примерно следующую схему:
Иногда в клетке образуются и третичные посредники. Так, обычно ионы Ca2+ выступают в роли вторичного посредника, но при передаче сигнала с помощью инозитолтрифосфата (вторичный посредник) выделяющиеся при его участии из ЭПР ионы Ca2+ служат третичным посредником.

